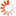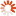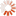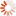
‘FREEDOM’—Chapter 4 The Meaning of Life
Chapter 4:5 Elaborating the sexually reproducing individual
As stated, while sexually reproducing individuals cannot normally be integrated, the sexually reproducing individual itself could be elaborated, made bigger, which, as will now be explained, is how single-celled organisms developed into multicellular organisms, and how multicellular colonial ants and bees integrated into their fully cooperative colonies. Struggling to find a way to develop greater order of matter by integrating sexually reproducing individuals, it was as if Negative Entropy (or God) decided, ‘Well, what I’ll do is develop greater order of matter within each genetically reproducing individual, making it bigger.’
As was also mentioned, in these ‘elaborated sexually reproducing individuals’, the cells of the multicellular body, or the individual ants and bees in their fully integrated colonies, are no longer sexually reproducing individuals themselves, but part of a larger sexually reproducing individual, which is the body, or, in the case of ants and bees, the colony.
The biological mechanism for elaborating the sexually reproducing individual involved the body’s cells, or the colony’s multicellular bees/ants, delegating the task of sexual reproduction to a distinct part of the whole that specialises in reproduction. In the case of the integration of single-celled organisms, the green alga known as Volvox provides an example of an organism in transition from the single-celled to the multicellular state, as this quote describes: ‘Volvox is…a small, green sphere…composed of thousands of flagellates embedded in the surface of a jelly ball…Volvox is a colony of unicellular animals rather than a many-celled animal, because even the simplest many-celled animals have considerably more differentiation between cells than appears among the cells of Volvox. The colony swims about, rolling over and over from the action of the flagella; but, remarkably enough, the same end of the sphere is always directed forward…Its behaviour can be explained only by supposing that the activities of the numerous flagellates are subordinated to the activity of the colony as a whole. If the flagella of each member of the colony were to beat without reference to the other members, the sphere would never get anywhere. In such subordination of the individual cells of a colony to the good of the colony as a whole we see the beginnings of individuality as it exists in the higher animals, where each animal behaves as a single individual, although composed of millions of cells…The co-ordination of numerous components into an individual is usually followed by the specialisation of different individuals for different duties. Only the slightest degree of specialisation is seen in the Volvox colony; the flagellates of the back part of the colony are capable of reproduction, while the front members never reproduce but have larger eyespots and serve primarily in directing the course of the colony’ (R. Buchsbaum et al., Animals without Backbones, 1938, p.50 of 401).
The marine invertebrates known as siphonophores, which include species such as the Portuguese man-of-war (otherwise known as the Bluebottle), live in colonies composed of ‘zooids’, individual animals that are not fully independent—indeed, their reliance upon and integration with each other is so strong the colony attains the character of one large organism. In fact, most of the zooids are so specialised they lack the ability to survive on their own. Thus siphonophorae, like Volvox, exist at the boundary between colonial and complex multicellular organisms.
We can imagine the path to the creation of Volvox and siphonophores began with cloning, which is the asexual reproduction of identical offspring where competition between the clones is pointless and unnecessary since each individual is genetically identical and, therefore, the division of labour and cooperation can develop and exist between individuals. One concern with cloning, however, is loss of variability—for example, if one colony kept reproducing asexually it could become so big it monopolised the available resources of food, space and territory, leading to the detriment of other colonies and a subsequent lack of variability in the species. We can imagine that eventually a limited, functional size would be arrived at through natural selection, which presumably is the size at which Volvox and siphonophores operate. And obviously to maintain variability, it would also be beneficial for sexual reproduction to occur from time to time, as it does amongst Volvox and siphonophores.
In the case of bees, the queen bee feeds all of her offspring that she intends to be workers a ‘royal jelly’ that causes sterility (ants also employ a similar chemical retardant). To ensure the reproduction of their genes these sterile offspring then have to support the queen because she carries their genes. (It should be mentioned that saying the queen ‘intends’ and the offspring ‘have to’ is obviously personifying the genetic process. The queen and the offspring are obviously not conscious thinking organisms, deciding they ‘intend’ and ‘have to’ do something or other as humans do; however, this form of anthropomorphism is simply a useful way of describing what, in effect, occurs. For example, the way genetics actually causes offspring to ‘have to’ support the queen is that, out of the many different mutational varieties of offspring that appear over time, only those that happen to have a genetic make-up that inclines them to support the queen will tend to reproduce, naturally selecting that particular behaviour for all subsequent generations and eventually the whole species.)
Elaborating the sexually reproducing individual allows the members of the elaborated individual to develop the ability to at least behave unconditionally selflessly, which, as has been explained, is fundamental if the fully cooperative integration of members into a new whole is to develop. The reason our body works so well is because each part has sublimated its needs to the greater good of the whole body; each part behaves unconditionally selflessly. Just as our skin cells are in constant turnover, with new cells replacing the old ones that have sacrificed themselves to protect our body, the leaves that fall in autumn do so to ensure their tree survives through winter. Bees and ants readily sacrifice themselves for their colony; for example, when a bee stings to protect its hive, its innards are attached to the sting that is left in its victim, so when it stings, it dies. The skin, leaves and bees/ants have behaved unconditionally selflessly; they have, in effect, considered the welfare of the greater good above their own welfare.
Of significance, however, is the emphasis here on our body’s skin, the tree’s leaves and the bees/ants only behaving unconditionally selflessly, because the selflessness apparent in these examples is not actually true unconditional selflessness, it is not true altruism. This is because the self-sacrificing skin, leaves and bees/ants are all indirectly selfishly ensuring their own genetic existence will be maintained by supporting the body, tree, or bee/ant colony that carries the genes for their existence and so reproduces them when it reproduces itself as a whole. Genetically, they are selflessly fostering the body/tree/colony to selfishly ensure their own genetic reproduction. Their apparently unconditionally selfless behaviour is not actually unconditional and thus altruistic, but rather a subtle form of selfishness. As explained earlier, such reciprocity can develop genetically because it doesn’t compromise the chances of the sexually reproducing individual reproducing its genes. (As pointed out in par. 197, the dishonest biological theory of Sociobiology/Evolutionary Psychology was truthful to the extent that it did recognise this fact that the selfless behaviour of social ants and bees is due to reciprocity—where the theory was dishonest was in its application of ‘kin selection’ to explain all social behaviour, even our own unconditionally selfless, universally benevolent, fully altruistic moral instincts.)
It now needs to be explained that large animals couldn’t employ this device of elaborating the sexually reproducing individual to develop a fully cooperative, integrated association or whole of their members because for them it involves too great a loss of the variability that all species need to be able to adapt to their environment. For example, if a female buffalo happened to be born with a particular mutation that caused her to produce a chemical in her milk that retarded the sexual maturation of most of her offspring such that those offspring then had to have selected mutations that inclined them to protect her to ensure their genes are successfully reproduced by her, and this became a common practice amongst buffalos with every queen buffalo having, say, 9 protector sacrificial buffalos from 10 offspring (so there is a sexual offspring to ensure the reproduction of the buffalo species, like ants have a few fertile females and males to carry on their species, but these fertile offspring have the same potential to produce some infertile offspring), then the genetic variety of a population of 1,000 buffalos would be reduced to just 100, a drastic loss of the variability so critical to ensuring that the species’ genetic pool remains able to adapt to any changes in its environment and thus keep maintaining and/or developing greater order. In the case of bees/ants, they are so small in relation to their environment that they can afford to have many fully integrated colonies in their environment without any significant loss of variability within their species.
The following two photographs illustrate the point. As will be mentioned shortly, while termites are a variety of cockroach rather than ant or bee, they have developed the same colonial capability as colonial ants and bees, which means that although there are millions of termites in each termite mound, in terms of the genetic variety present in the territory shown, these mounds do, in fact, represent a similar number of sexually reproducing individuals to the number of sexually reproducing individual buffalos shown in a corresponding area in the second photograph.

Magnetic Termite Mounds, Litchfield National Park, Northern Territory, Australia;
and feral Asian Buffalo, Northern Territory, Australia. Photographs by the author, 2010.
Quite a number of species that are much larger than ants and bees are attempting to create the integrated society of members by temporarily elaborating the sexually reproducing individual. Many bird species, such as the Australian kookaburra, delay their sexual maturation for a few years after they fledge, during which time they selflessly help raise their parents’ subsequent offspring. Wolves, African wild dogs and meerkats do the same thing. However, what these species have obviously found is that to delay their sexual maturation permanently leads to too great a loss of variability in their species.
Underground-living colonial naked mole rats form fully integrated colonies of up to 300 members comprising a single queen who uses hormones to inhibit the sexual maturation of nearly all the others who then act as ‘workers’ and ‘soldiers’. A few ‘sexual disperser caste’ are allowed to reach sexual maturity and these periodically leave their natal burrow to access other colonies and, in doing so, help maintain the genetic variety of the mole rat species. Significantly, like colonial ants and bees, and the dozen or so other varieties of multicellular organisms (including the termite) that have been able to permanently elaborate the sexually reproducing individual, mole rats are relatively small; individuals typically measure only 8 to 10 centimetres (3 to 4 inches) long.
What has been explained here is very significant for humans because it means, as large animals, we could not have employed the integrating device of elaborating the sexually reproducing individual to create the pre-conscious and pre-human-condition-afflicted, fully cooperative, completely integrated, ‘Golden’, ‘Garden of Eden’-like state that our distant ancestors lived in. Further, during that fully integrated, idyllic past our instinctive orientation was not to reciprocity’s subtle form of selfishness that the parts of multicellular organisms and bee/ant colonies practise, as the theory of Sociobiology/Evolutionary Psychology claims, but to being truly altruistic, genuinely unconditionally selflessly orientated towards all of life. Thus, even if we could have employed the device of elaborating the sexually reproducing individual it would not even begin to account for our unconditionally selfless moral soul. I italicised ‘all of life’ because while ant and bee colonies have members who are dedicated to supporting each other, each colony is, in fact, engaged in fierce competition with other colonies. Worker ants and bees are not interested in behaving selflessly towards all of life, which, contrary to what the theory of Multilevel Selection claims, our moral self is interested in. As pointed out in chapter 2:11, our ability to love unconditionally didn’t arise from an ability to war successfully. So, the question remains: how did humans manage to develop our absolutely wonderful and astonishing unconditionally selfless, genuinely altruistic, all-loving moral instinctive orientation to the world?